4. Discussion
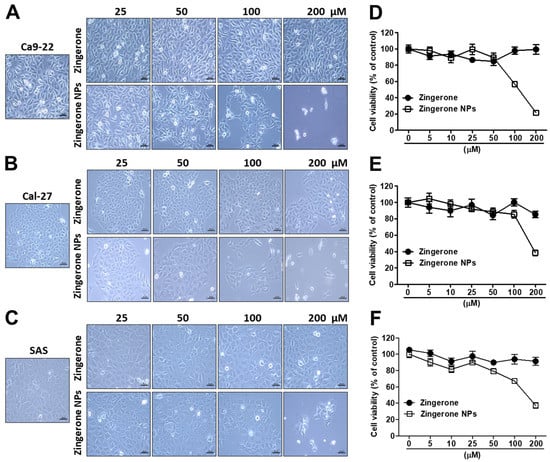
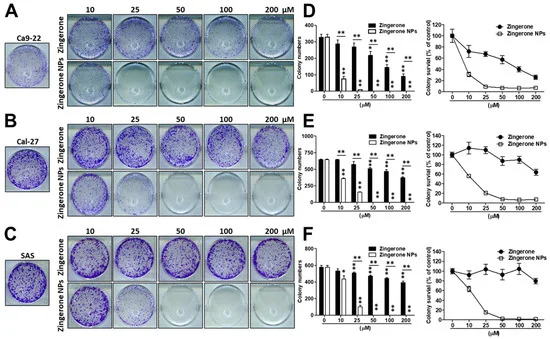
Previous studies have indicated that zingerone exhibits different cytotoxic effects on different cancer cell types. For example, a lower dose (IC50 < 25 µM) of zingerone treatment has been shown to induce >50% cytotoxicity in both colon cancer and prostate cancer [40,41], while a high dose (IC50 > 2000 µM) of zingerone treatment merely elicited 40% cytotoxicity in neuroblastoma cells [42]. Indeed, higher tolerance to zingerone treatment (IC50 is over 200 µM) was also observed in OSCC cells and hepatoma cells [29]. Here, we fabricated green and carbon-based zingerone NPs and showed that they possessed superior cytotoxicity and antitumorigenic activity. For instance, a lower dose (25 µM) of zingerone NP treatment inhibited tumorigenesis by 50% and 80% in hepatoma cells [29] and OSCC cells, respectively, whereas higher doses of zingerone (>200 µM) may be needed to achieve a similar efficacy. Notably, the safety of zingerone NPs was also validated in our previous study. After mice were fed either zingerone or zingerone NPs for 8 weeks (0.4 mg/25 g BW), neither treatment elicited significant liver toxicity (based on the GOT/GPT index of the sera) [29]. Moreover, in vitro data also validated that the normal bone marrow HS-5 cells had higher tolerance to zingerone NP treatment (IC50 = 184.2 ± 6.8 µM) compared to these OSCC cell lines (Figure S4). This result suggested that zingerone NPs revealed good biocompatibility and safety toward normal cells.
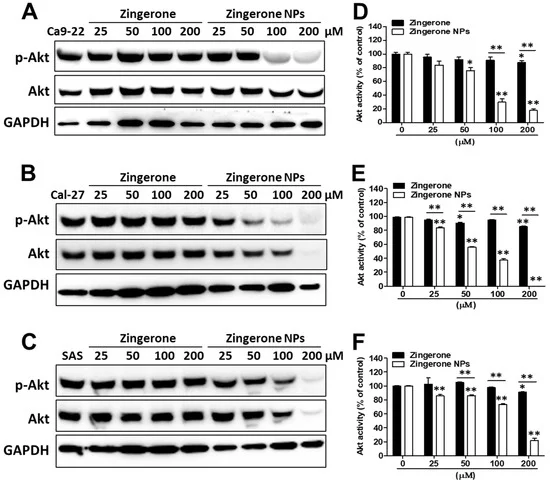
The as-prepared zingerone NPs had an ultrasmall size of 1.42 ± 0.67 nm and were clear with an amber color. Moreover, zingerone NPs have a negative charge (−15.99 ± 0.23 mV), and an absorption peak at 350 nm is observed in the UV–vis spectrum due to the π−π* transition of the benzenoid rings. The total ion chromatogram analysis also reveals functional groups, including methoxy and hydroxyl groups, on the nanoparticle. Alterations in the physicochemical properties may increase the pharmacological efficacy of zingerone NPs in cancer cells. Indeed, nanosized zingerone may have more surface chemistry consisting of methoxy and hydroxyl groups than zingerone alone [29]. Moreover, methoxy groups and hydroxyl groups have been reported play pivotal roles in the bioactivities of natural compounds, such as anticancer, antimigration, and antiproliferative activities [43]. Furthermore, unlike positively charged nanoparticles, which are internalized by cells through an electrostatic interaction between cationic NPs and the negatively charged cell membrane, negatively charged nanoparticles adsorb to the nuclear membrane and interact with nuclei, potentially because the nucleolar pH is consistently 0.3 to 0.5 units higher than that of the cytosol [44,45,46]. Therefore, variations in physicochemical properties, such as the ultrasmall size, shape, surface charge, and surface chemistry, may increase the uptake of nanoparticles by cells and allow them to interact with macromolecules, such as nucleotides, lipids, and proteins, thus interfering with cellular biofunctions [47,48]. These outcomes may ultimately induce lethal stress in cancer cells. On the other hand, we have reported that zingerone NPs significantly increased cell apoptosis, decreased cell proliferation and disturbed the cell cycle distribution in human hepatoma cells. These results are attributed to the zingerone NP-mediated downregulation of Akt/NF-κB signaling, increased DNA damage and DNA instability, and the activation of caspase-mediated apoptosis signaling pathways. These outcomes finally contribute to the inhibitory effects of zingerone NPs on the proliferation and tumorigenicity of hepatoma cells [29]. Accordingly, both Akt/NF-κB signaling and the caspase signaling cascade might be involved in the zingerone NP-mediated inhibitory effects on proliferation and tumorigenicity in OSCC cell lines.
Recently, zingerone has attracted increasing interest from researchers for its chemopreventive potential, including eliciting antiangiogenic activity to protect against tumor development [49], protecting against colon carcinogenesis mediated by the carcinogen 1,2-dimethylhydrazine (DMH) in rats [50], and inducing the mitotic arrest of the cell cycle to suppress neuroblastoma development [42]. However, a relatively higher dose is necessary to achieve therapeutic efficacy. To date, compared to the raw phytochemical compounds, phytochemically derived carbon dots (CDs) have been reported to have superior efficacy in biomedical applications, including antiviral activity and anticancer efficiency. For instance, curcumin-derived carbon quantum dots (Cur-CQDs) significantly inhibited enterovirus 71 (EV71) infection (half-maximal effective concentration (EC50) < 20 µg/mL compared to curcumin (EC50 > 200 µg/mL)) and protected newborn mice from virus-induced hindlimb paralysis [51]. Moreover, berberine is an isoquinoline alkaloid that possesses good anticancer, antibacterial, and anti-inflammatory bioactivities [52]. A previous study reported that berberine-based carbon dots (Ber-CDs) not only had superior optical properties for bioimaging and retained the biofunctions of berberine, but also exhibited selective and safe antitumor performance (with a decrease of approximately 50% of tumor nodules) [53]. In the present study, a lower dose (<25 µM) of zingerone-based carbon dots (zingerone NPs) not only elicited significant antiproliferative and antitumor activity, but also inhibited the migration and invasion of OSCC cells.
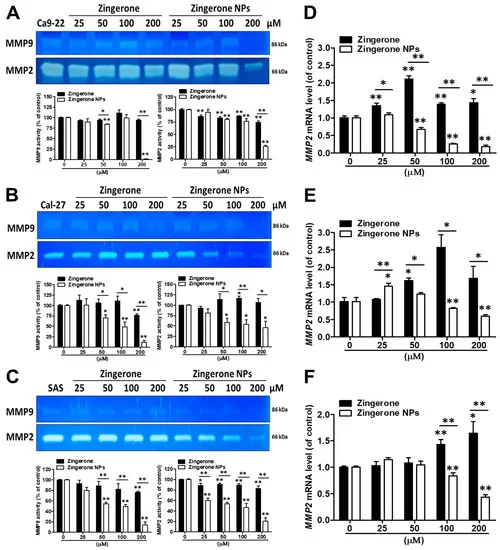
Cancer cells that metastasize into the cervical lymph nodes and/or distant organs have become the most important prognostic indicator of oral squamous cell carcinoma (OSCC). Moreover, several signaling mechanisms have been identified that are involved in OSCC invasion and metastasis, such as MMP activity, cadherin switching-associated EMT-related signaling cascades, and signaling pathways, such as the PI3K/Akt signaling, TGF-β, and Wnt/β-catenin pathways [54]. Based on accumulating evidence, phytochemicals interfere with various intracellular mechanisms in many types of cancer, including the induction of apoptosis, the arrest of cell cycle progression, the blockade of transcription factors, such as NF-κB, the induction of oxidative stress, the suppression of MMP activity, and the inhibition of signal transduction pathways, such as the Akt, VEGF, Wnt, and STAT3 signaling pathways [21,55]. In addition, as potent adjuvants, accumulating studies have suggested that phytochemicals significantly improve cancer invasion and metastasis, and attenuate side effects and drug resistance during treatment with conventional tumor therapeutics [56,57].
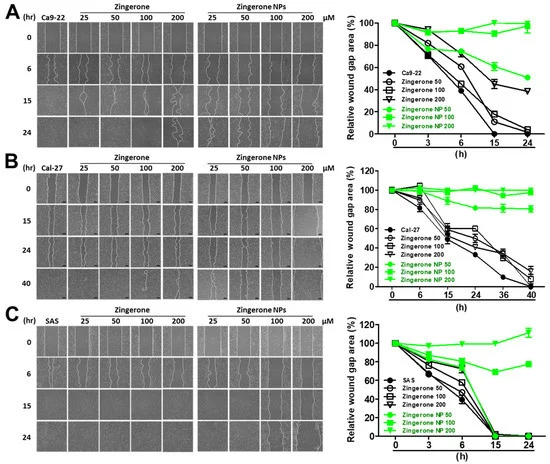
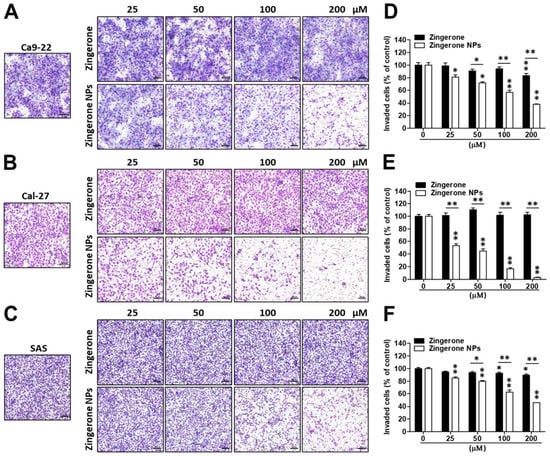
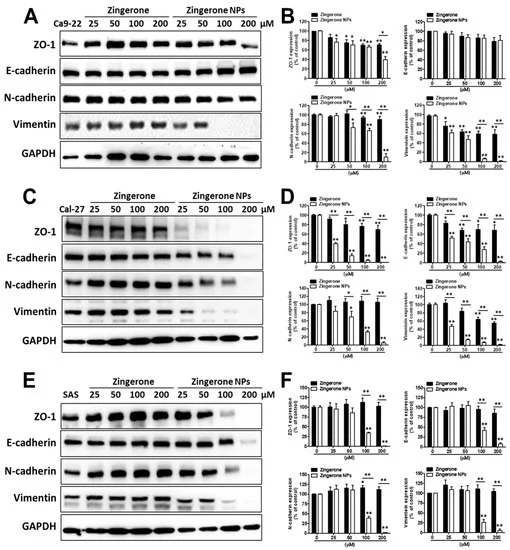
Additionally, compared to the tongue squamous cell carcinoma cell lines, Cal-27 and SAS, zingerone NPs showed greater bioactivity and suppression of gingival squamous cell carcinoma Ca9-22 cells activity in the MTT assay (Figure 1), colony formation assay (Figure 2), wound healing assay (Figure 3), and MMP activity assay (Figure 6). Interestingly, by analyzing the EMT-related markers, zingerone NPs obviously reduced the levels of epithelial markers in the Cal-27 and SAS cell lines rather than in Ca9-22 cells (Figure 7). Zingerone NPs also exerted different inhibitory effects on invasion (Figure 4) and Akt activity (Figure 5) among these OSCC cell lines. Although the Ca9-22, Cal-27, and SAS cell lines were classified as aggressive OSCC cells in the cancer grading system, differences in the genome and/or gene diversity, and variations in the cellular molecular biology, may guide different cellular responses to environmental stimuli. For example, Ca9-22 cells, a malignant gingival squamous cell carcinoma line that was extracted from a 43-year-old Japanese male, were identified as tetraploid cells without the typical normal chromosome form 46XY [58]. Cal-27 cells are a form of malignant tongue squamous cell carcinoma that were extracted from a 56 year-old Caucasian male, and a cytogenetic analysis revealed moderate hyperploidy; the average number of chromosomes per cell analyzed was 43. One of the chromosomes of certain pairs was always (pairs 8, 9, 11, 13, 22) or almost always (pairs 12, 21) missing [59]. The SAS cells, representing poorly differentiated tongue squamous cell carcinoma, were extracted from a 69-year-old Japanese female [60]. All the different genomic backgrounds may contribute to the differences in tolerance to zingerone and/or zingerone NP treatment. Moreover, the gene expression profile and differences in genetic heterogeneity in different races and sexes would also confer differences in sensitivity to zingerone and/or zingerone NP treatment.
Altogether, we combined both phytochemicals and nanotechnology to enhance raw zingerone bioactivity, and attempted to develop zingerone NPs as potent nanomedicines and/or phytochemical nano-adjuvants for not only hepatocellular carcinoma cells [29], but also oral squamous carcinoma cells. In this study, the as-prepared zingerone NPs exhibited impressive efficacy in decreasing levels of cell invasion-associated factors, such as MMPs and Akt signaling, and interfered with EMT-associated proteins, which all led to reduced cell survival and inhibited the invasion and metastasis of OSCC cells (Figure 8).
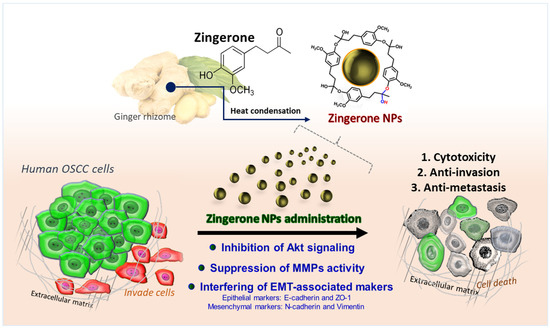
Figure 8. The phytochemically derived zingerone NPs elicited superior efficacy to inhibit the cell invasion and metastasis of human oral squamous cell carcinoma. The as-fabricated zingerone NPs significantly suppressed Akt signaling-mediated cell survival and cell motility, which led to obvious cytotoxicity and anti-proliferation. Moreover, the zingerone NP-mediated downregulation of MMP activity was also reflected in the harsh cell motility. In addition, the zingerone NPs substantially disturbed the expression levels of EMT-associated markers, including the mesenchymal markers N-cadherin and vimentin, and the epithelial markers ZO-1 and E-cadherin. These results suggested that the zingerone NPs exerted superior suppression on cell proliferation, tumorigenicity, and cell motility, and thus achieved an inhibitory effect against the cell invasion and metastasis of human oral squamous cell carcinoma.
5. Conclusions
Nanoscience is rapidly reorganizing the landscapes of various cancer therapeutic strategies and chemopreventive agents. In the present study, we fabricated phytochemically derived zingerone NPs and documented their superior bioactivity and multiple pharmacological effects on OSCC cells, including their inhibitory effects on proliferation, motility, invasion, and tumorigenesis. Our data indicate that zingerone NPs significantly suppress the activities of the Akt protein and MMP enzymes, decrease MMP2 mRNA expression, and disturb EMT marker expression. Zingerone NPs thus exert potent inhibitory effects on the invasion and metastasis of OSCC cells.
Altogether, these results suggest that zingerone NPs may represent a potent chemopreventive adjuvant and provide an alternative strategy for treating OSCC that improves the side effects, drug resistance, and recurrence associated with conventional cancer therapies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biomedicines10020320/s1, Figure S1: Characterization of zingerone NPs reproducibility; Figure S2: Cytotoxicity effects of Zingerone NPs at short-term incubation of OSCC cell lines; Figure S3: Effect of zingerone NPs on the enzyme activity of lower molecular weight MMPs in human OSCC cell lines; Figure S4: Effect of Zingerone NPs on cytotoxicity of normal human bone marrow HS-5 cells.
Author Contributions
Conceptualization, S.H. and M.-L.K.; Data curation, C.-M.Y., T.-H.C. and K.-W.T.; Formal analysis, K.-W.T.; Investigation, C.-M.Y., T.-H.C., K.-W.T., S.H. and M.-L.K.; Methodology, T.-H.C., S.H. and M.-L.K.; Project administration, C.-M.Y.; Supervision, M.-L.K.; Validation, T.-H.C.; Writing–original draft, C.-M.Y. and M.-L.K.; Writing–review & editing, C.-M.Y. and M.-L.K. All authors have read and agreed to the published version of the manuscript.
Funding
Grants were received from the Ministry of Science and Technology of Taiwan (MOST 110-2314-B-075B-004-MY3), and Kaohsiung Veterans General Hospital (KSVGH110-098 and VGHKS109-177).